The Four Intertwined Principles of Multicelularity
In biology the four principles -- specialization, messaging, stigmergy and apoptosis -- evolved together and depend upon each other. They should do so in multicellular computing as well.
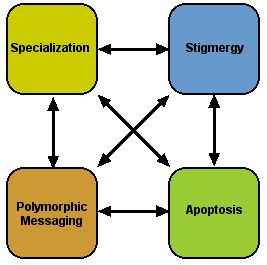
Each cell participates simultaneously in all four principles
In multicellular organisms each differentiated cell functions in a specialized way as part of a coherent larger system that evolved as a whole. That is, the specializations coevolved -- one specialization supports and depends upon another. Similarly, the four principles coevolved during this process so that virtually all cells participate in all four architectural realms at once.
 Specialization
-- All healthy Metazoan cells are specialized. Even adult stem cells
are
somewhat specialized. What is perhaps the
most specialized aspect of the cells, other than their unique shape and
function, is their unique repertoire of message receptors that
determine
the set of message molecules to which they can respond.
They all share common behaviors too. Included in the common
behavior are participation in the cues and signals of their stigmergy
relationship with the rest of the body, and obedience to apoptosis
messages. That is, as multicellular organisms evolved specialized
behaviors, they had to also evolve appropriate messaging, stigmergy and
apoptosis behaviors.
Specialization
-- All healthy Metazoan cells are specialized. Even adult stem cells
are
somewhat specialized. What is perhaps the
most specialized aspect of the cells, other than their unique shape and
function, is their unique repertoire of message receptors that
determine
the set of message molecules to which they can respond.
They all share common behaviors too. Included in the common
behavior are participation in the cues and signals of their stigmergy
relationship with the rest of the body, and obedience to apoptosis
messages. That is, as multicellular organisms evolved specialized
behaviors, they had to also evolve appropriate messaging, stigmergy and
apoptosis behaviors.
Polymorphic Messaging -- Complex messenger proteins often act as "bundles" of messages. That is, one messenger protein may have separate domains, each with a different messaging function [1]. And often, the different message domains address each of the other three architectural principles. For example, one domain initiates signal cascades specific to the unique specialized function of that type of cell, another domain on the same complex molecular messenger facilitates or verifies physical attachment to the extracellular matrix (i.e., deals explicitly with the stigmergy structure), and yet another provides signals that either suppress or encourage apoptosis! The existence of these multi-part messages shows not only that the organisms evolved along with the four principles, but also how fundamental these principles are. A single multi-part message speaks to the functional relationship of the cell to the whole organism/tissue/organ rather than to just a single cell function.
Stigmergy -- Virtually all cells other than cells such as red blood cells are affected by stigmergy cues and/or signals. Even unattached cells such as blood and lymph born cells are affected by and affect blood borne stigmergy signals, e.g. hormones. Cells that are attached to the Extracellular Matrix (ECM), i.e., the stigmergy structure, leave long-lasting cues (persistent messages) in those structures that affect other cells. In turn, the cells respond to such cues in ways that may cause them to modify the physical structures; that's how the structures are built in the first place. Cells that are normally attached or in direct contact with the ECM require constant feedback from the ECM. Absent the appropriate attachment cues, they suicide (undergo apoptosis).
Apoptosis -- almost all cells except cancerous cells participate in apoptosis signaling all the time. Even very simple cells such as red blood cells that lack a nucleus undergo apoptosis.
The principles themselves are operationally interdependent
Not only do cells reflect all four principles at once, the principles themselves are interdependent in the sense that each one relies on the others.
- Specialization requires a stigmergy structure (a body)
to nurture and protect the specialized cells. They would
not survive long in isolation. In turn, the stigmergy structure, i.e.,
the whole organism, benefits from
their specialized activity. A stigmergic body makes
no sense without specialization; Even cells in biofilms specialize.
So, too, do ants and termites. The
more complex their stigmergic social interactions, the more specialized
are their roles in the insect colony (see pdf).
Thus
specialization and
stigmergy are
interdependent at a fundamental biological level.
- Once cells specialize, they must interpret messages
accordingly, i.e., polymorphicly. That's how a collaborating
group with various
specializations can respond in an orchestrated way to common
stimuli. There is no orchestra conductor telling each one what
to do. A common message signals the situation and each
specialized cell with a receptor for that message responds in
accordance with its specialized role. Similarly, specialized ants
and termites must interpret
messages from the perspective of their special abilities. So
specialization and polymorphic messaging are strongly interdependent.
- Since apoptosis exists to sculpt and protect a (stigmergic) body, clearly stigmergy and apoptosis are interdependent. But also, the apoptotic messaging pathways depend upon polymorphic messaging and the cellular response to apoptosis messages differs according to the specialized function of the cell. Thus apoptosis and specialization are interdependent.
Implications for multicellular computing
As multicellular computing architectures evolve, especially the emerging Service Oriented Architectures (SOA) and less formal Web Service mashups, we would be wise to adopt and carefully interleave all four principles. Similarly, the architecture of multicellular computing messages should also be fully multicellular. Those who "design" multicellular computing systems (or better put, attempt to grow and evolve such systems), should not only give thought to the various kinds of specialized "cells" that are needed, but also to how those cells implement the four principles and fit into a multicellular message architecture.
In summary, as we develop architectural patterns for the use of each of the four principles in both hardware and software, we must develop them so that the four patterns can be interwoven. And there must be a meta-pattern that lays out how the interweaving is to happen.
[1] See "Exploring and Engineering the Cell Surface Interface" abstract (Stevens, M. M. & George, J. H., Science, vol 310, Nov. 18, 2005, pp. 1135-1138)
Contact: sburbeck at mindspring.com
Last revised 6/9/2012